
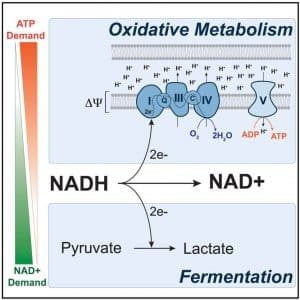
 Nearly a century ago, Otto Warburg discovered that tumors consume tremendous amounts of glucose relative to most non-transformed tissues, and that the majority of glucose consumed by tumors is fermented to lactate, rather than oxidized in pathways that require respiration. This phenotype is referred to as “aerobic glycolysis,” because unlike carbohydrate fermentation in response to oxygen limitation, aerobic glycolysis involves high levels of fermentation even when oxygen is abundant. Aerobic glycolysis a hallmark of proliferative metabolism found across many kingdoms of life, but is frequently associated with cancer cells, and is known as the Warburg effect in this context.
Nearly a century ago, Otto Warburg discovered that tumors consume tremendous amounts of glucose relative to most non-transformed tissues, and that the majority of glucose consumed by tumors is fermented to lactate, rather than oxidized in pathways that require respiration. This phenotype is referred to as “aerobic glycolysis,” because unlike carbohydrate fermentation in response to oxygen limitation, aerobic glycolysis involves high levels of fermentation even when oxygen is abundant. Aerobic glycolysis a hallmark of proliferative metabolism found across many kingdoms of life, but is frequently associated with cancer cells, and is known as the Warburg effect in this context.
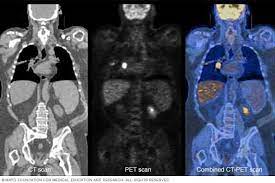 Elevated glucose uptake by tumors is detected by fluorodeoxyglucose-positron emission tomography (FDG-PET) imaging for initial cancer staging, assessing response to therapy, and surveillance. Beginning with the initial observation by Otto Warburg and others nearly a century ago that tumor cells increase glucose uptake and produce high quantities of lactate, even in the presence of oxygen, it has been well established that cancer cells engage in altered metabolism.
Elevated glucose uptake by tumors is detected by fluorodeoxyglucose-positron emission tomography (FDG-PET) imaging for initial cancer staging, assessing response to therapy, and surveillance. Beginning with the initial observation by Otto Warburg and others nearly a century ago that tumor cells increase glucose uptake and produce high quantities of lactate, even in the presence of oxygen, it has been well established that cancer cells engage in altered metabolism.
 Glucose is the single most abundant nutrient for most cells and can be a source of biomass and fuel for energy production. Numerous signaling pathways altered in cancer affect glucose metabolism through a variety of mechanisms. In one classic paradigm, receptor tyrosine kinases induced by insulin or other growth factors activate the PI3K-AKT signaling pathway to stimulate glycolysis.
Glucose is the single most abundant nutrient for most cells and can be a source of biomass and fuel for energy production. Numerous signaling pathways altered in cancer affect glucose metabolism through a variety of mechanisms. In one classic paradigm, receptor tyrosine kinases induced by insulin or other growth factors activate the PI3K-AKT signaling pathway to stimulate glycolysis.
 Compared with most normal tissues, cancer cells have an increased demand for fatty acids to generate lipid membranes and precursors for signaling molecules. Fatty acids can be acquired exogenously through diet or synthesized endogenously from glucose, glutamine, or acetate.
Compared with most normal tissues, cancer cells have an increased demand for fatty acids to generate lipid membranes and precursors for signaling molecules. Fatty acids can be acquired exogenously through diet or synthesized endogenously from glucose, glutamine, or acetate.
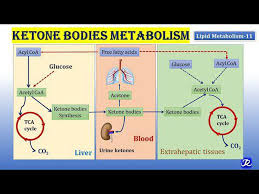 Some cancers lack the ability to metabolize ketone bodies, due to mitochondrial dysfunction and down-regulation of enzymes necessary for ketone utilization. A fat-rich, low-carbohydrate diet in cancer therapy aims to reduce circulating glucose levels and induce ketosis such that cancer cells are starved of energy while normal cells adapt their metabolism to use ketone bodies and survive. Furthermore, by reducing blood glucose also levels of insulin and insulin-like growth factor, which are important drivers of cancer cell proliferation, drop. Numerous preclinical studies have provided evidence for an anti-tumor effect of KDs.
Some cancers lack the ability to metabolize ketone bodies, due to mitochondrial dysfunction and down-regulation of enzymes necessary for ketone utilization. A fat-rich, low-carbohydrate diet in cancer therapy aims to reduce circulating glucose levels and induce ketosis such that cancer cells are starved of energy while normal cells adapt their metabolism to use ketone bodies and survive. Furthermore, by reducing blood glucose also levels of insulin and insulin-like growth factor, which are important drivers of cancer cell proliferation, drop. Numerous preclinical studies have provided evidence for an anti-tumor effect of KDs.